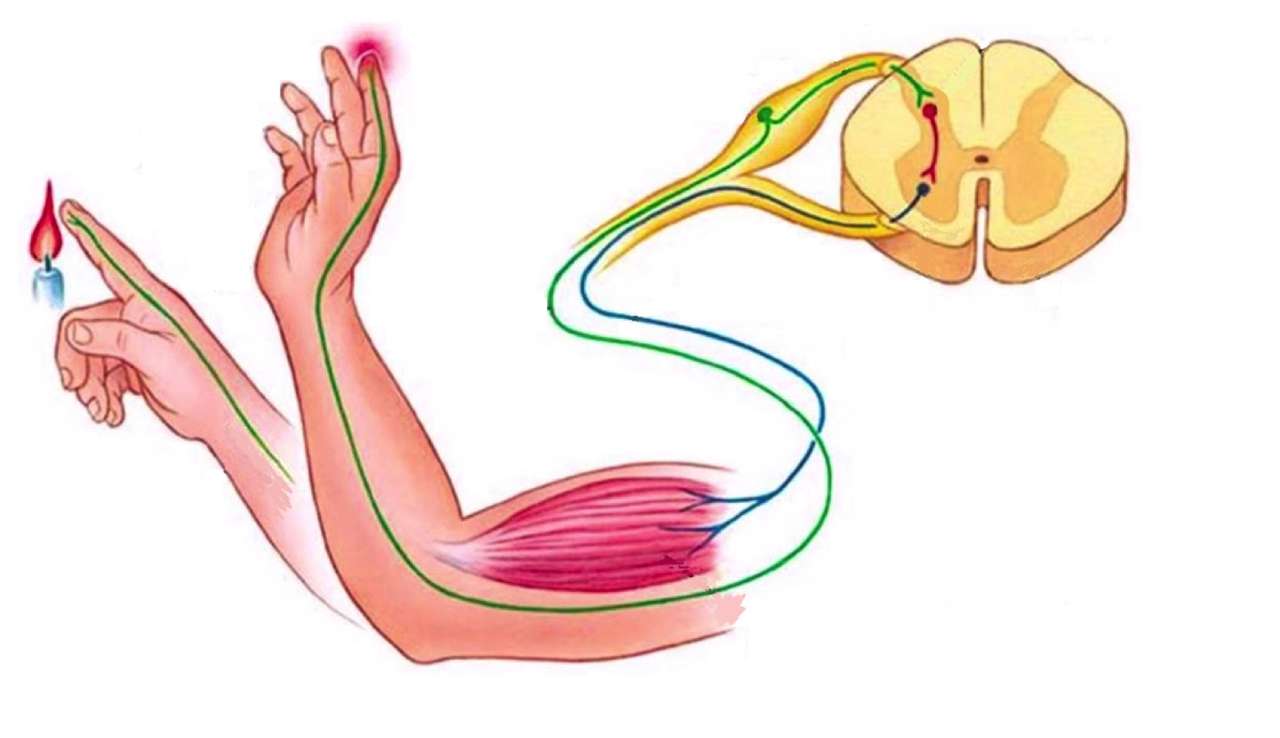
Рефлекторна дуга є основоположною структурою нервової системи людини та тварин, яка перетворює подразнення на автоматичну рухову або секреторну відповідь, минаючи свідомий аналіз у головному мозку. Цей механізм забезпечує захист від пошкоджень, підтримку пози тіла та регуляцію внутрішніх органів з мінімальною затримкою в часі.
Її анатомічна організація включає послідовність з п’яти елементів, де кожен відіграє критичну роль у проведенні нервового імпульсу та його трансформації. Порушення в будь-якій ланці цієї дуги може призвести до серйозних неврологічних порушень, що робить її вивчення важливим як для базового розуміння фізіології, так і для клінічної практики.
У глибшому розгляді рефлекторна дуга демонструє елегантність еволюційних рішень: від простих моносинаптичних ланцюгів до складних полісинаптичних мереж з гальмуванням та полегшенням, що дозволяє адаптивні та координовані реакції організму.
П’ять ключових компонентів рефлекторної дуги
Рефлекторна дуга функціонує як чітко організований ланцюг, де кожна ланка виконує специфічне завдання. Початок завжди лежить у рецепторі — спеціалізованому утворенні, яке перетворює енергію подразника (механічну, термічну, хімічну чи больову) на електричний сигнал. Рецептори розташовані в шкірі, м’язах, суглобах, внутрішніх органах та органах чуття. Вони генерують градуйований рецепторний потенціал, сила якого залежить від інтенсивності подразнення. Якщо сигнал досягає порогу, виникає потенціал дії.
Далі імпульс прямує по аферентному (чутливому) нейрону. Ці нейрони найчастіше псевдоуніполярні: їхні тіла лежать у спинномозкових вузлах, а довгі відростки проводять сигнал від периферії до спинного мозку через задні корінці. Для швидких рефлексів, таких як сухожильні, використовуються товсті мієлінізовані волокна групи Ia, що забезпечують високу швидкість проведення — до 120 м/с. Закон Белла-Мажанді чітко розмежовує: задні корінці — чутливі, передні — рухові.
У нервовому центрі відбувається обробка інформації. Для спінальних рефлексів це сіра речовина спинного мозку. Тут сигнал переходить через синапси: збуджувальні (з вивільненням глутамату та генерацією збуджувального постсинаптичного потенціалу) або гальмівні (з ГАМК та гальмівним постсинаптичним потенціалом). У простих випадках центр умовний — пряма передача, у складних — участь інтернейронів з просторовою та часовою сумацією, полегшенням чи оклюзією.
Еферентний (руховий) нейрон несе команду від центру до периферії. Його тіло розташоване в передніх рогах спинного мозку, аксон виходить через передні корінці та іннервує ефектор. Для скелетних м’язів це альфа-мотонейрони, які формують нервово-м’язовий синапс з ацетилхоліном.
Завершує ланцюг орган-ефектор — м’яз (скелетний, гладкий або серцевий) чи залоза. Тут імпульс перетворюється на дію: скорочення, розслаблення або секрецію. Градуйована відповідь м’яза залежить від частоти імпульсів та кількості активованих рухових одиниць.
Механізм передачі сигналу в рефлекторній дузі
Процес розпочинається з трансдукції: подразник змінює проникність мембрани рецептора, створюючи локальний струм. При досягненні порогу в аферентному волокні запускається потенціал дії — феномен «все або нічого». Імпульс швидко досягає спинного мозку або стовбура головного мозку.
У синапсі відбувається хімічна передача. Збуджувальні синапси деполяризують постсинаптичну мембрану, гальмівні — гіперполяризують. Сумація кількох сигналів вирішує, чи виникне потенціал дії в мотонейроні. У моносинаптичних дугах затримка мінімальна — лише 0,5–1 мс на синапс. Полісинаптичні додають час на обробку в інтернейронах, але дозволяють гнучкішу відповідь, включно з одночасним збудженням одних м’язів та гальмуванням антагоністів.
На рівні ефектора в нервово-м’язовому з’єднанні ацетилхолін відкриває іонні канали, викликаючи скорочення. У вегетативних дугах ефекторами стають гладкі м’язи чи залози з іншими медіаторами — норадреналіном чи ацетилхоліном залежно від відділу.
Важливо, що вищі відділи мозку можуть модулювати рефлекси: посилювати чи пригнічувати їх через низхідні шляхи. Це дозволяє свідомо контролювати деякі реакції, наприклад, не відсмикнути руку під час медичної процедури.
Різновиди рефлекторних дуг: від простих до складних
За кількістю синапсів розрізняють моносинаптичні та полісинаптичні дуги. Моносинаптичні містять лише один синапс між аферентним та еферентним нейроном — це найшвидші реакції. Полісинаптичні включають один або кілька інтернейронів, що дозволяє інтеграцію сигналів з різних джерел та координовані дії кількох м’язів.
| Характеристика | Моносинаптична дуга | Полісинаптична дуга |
|---|---|---|
| Кількість синапсів | Один | Два і більше |
| Швидкість реакції | Максимальна (латентність ~18–20 мс для колінного рефлексу) | Дещо повільніша, але з можливістю координації |
| Приклад | Колінний (надколінковий) рефлекс | Рефлекс відведення (withdrawal), перехресний розгинальний рефлекс |
| Складність обробки | Пряма передача, мінімальна інтеграція | Сумація, реципрокне гальмування, участь кількох сегментів спинного мозку |
За типом ефектора дуги поділяють на соматичні (скелетні м’язи) та вегетативні (гладкі м’язи, залози, серце). Соматичні рефлекси зазвичай спінальні або краніальні, вегетативні — часто з центрами в стовбурі мозку чи автономних гангліях. За місцем інтеграції виділяють спінальні (швидкі, захисні) та краніальні (наприклад, зіничний рефлекс на світло).
Класичні приклади рефлексів та їх особливості
Колінний рефлекс — класичний моносинаптичний приклад. Удар по сухожилку чотириголового м’яза розтягує м’язові веретена (пропріорецептори). Імпульси по Ia-волокнах досягають спинного мозку (сегменти L2–L4) і прямо збуджують альфа-мотонейрони, що іннервують чотириголовий м’яз. Одночасно через інтернейрон гальмується антагоніст — задній м’яз стегна (реципрокне гальмування). Це забезпечує плавне розгинання без протидії. Гамма-мотонейрони регулюють чутливість веретен, підтримуючи тонус м’язів.
Рефлекс відведення (флексорний, ноцицептивний) — полісинаптичний захисний механізм. Больовий подразник (укол, опік) активує ноцицептори, сигнал іде до кількох інтернейронів у спинному мозку. Результат — швидке згинання кінцівки та часто перехресне розгинання протилежної для збереження рівноваги. Така координація рятує від падіння під час раптового відсмикування.
Інші приклади: рефлекс моргання (захист ока від подразника), сухожильні рефлекси (ахіллів — S1–S2), вегетативні — барорефлекс (регуляція тиску крові) чи слиновиділення при запаху їжі. У немовлят присутні примітивні рефлекси (Моро, хватальний), які зникають із дозріванням кори головного мозку.
Клінічне значення рефлекторних дуг у сучасній медицині
Тестування глибоких сухожильних рефлексів — невід’ємна частина неврологічного обстеження вже понад століття. Лікарі використовують неврологічний молоточок для перевірки колінного, ахіллового, біцепсового, трицепсового та інших рефлексів. Градація від 0 (відсутній) до 4+ (з клонусом) допомагає локалізувати ураження.
Гіпо- або арефлексія вказує на ураження нижнього мотонейрона: периферичний нерв, корінець, передній ріг спинного мозку або нервово-м’язове з’єднання (поліневропатія, радикулопатія, СМА). Гіперрефлексія з патологічними знаками (Бабінського) свідчить про ураження верхнього мотонейрона — інсульт, розсіяний склероз, травма спинного мозку вище рівня сегмента. Асиметрія рефлексів часто першим сигналізує про локальну патологію.
Маневри підкріплення (наприклад, Джандрассіка) посилюють рефлекси при слабкій відповіді. У гострій стадії спінального шоку рефлекси тимчасово зникають навіть при повному перетині. Сучасна медицина поєднує класичне тестування з електронейроміографією для точнішої діагностики, проте рефлекторна дуга залишається доступним і надійним орієнтиром.
Цікаві факти про рефлекторні дуги
- Латентність колінного рефлексу становить лише близько 18–20 мс від моменту розтягнення сухожилка до початку скорочення м’яза — це в десятки разів швидше за свідомому реакцію (понад 200 мс).
- Сер Чарльз Шеррінгтон (Нобелівська премія 1932 року) не лише детально описав рефлекторні дуги, а й увів термін «синапс» та показав, що рефлекси — це інтегровані дії всього організму, а не ізольовані ланцюги.
- У полісинаптичних дугах одночасно може відбуватися реципрокне гальмування антагоністів та перехресне збудження — це дозволяє, наприклад, відсмикнути одну ногу та одночасно випрямити іншу для збереження рівноваги.
- Деякі рефлекси зберігаються навіть у стані глибокої коми чи під час сну, оскільки їхні центри розташовані в спинному мозку або стовбурі й не потребують участі кори.
- Гамма-мотонейрони, що іннервують інтрафузальні волокна м’язових веретен, підтримують постійну чутливість пропріорецепторів — без них рефлекси втрачали б точність при зміні довжини м’яза.
- У немовлят примітивні рефлекси (Моро, хватальний) зникають до 4–6 місяців саме через дозрівання гальмівних впливів кори головного мозку на спінальні дуги.
Розуміння рефлекторної дуги допомагає не лише лікарям у діагностиці, а й кожному — усвідомити, наскільки досконало влаштована наша нервова система для миттєвого захисту та підтримки життєдіяльності. Ця фундаментальна схема продовжує надихати нейрофізіологів і слугує основою для розробки нейропротезів та реабілітаційних технологій.